

The smallpox virus
We welcome our colleague Kyle Harper to the blog; his bio is on the OU contributors page. His book, The Fate of Rome: Climate, Disease, and the End of an Empire, is now available from Princeton University Press. This blog post expands from Harper’s work on the Roman Empire to look more broadly at the long history of interaction between human societies and pathogenic microbes.
In AD 166, the city of Rome was invaded by an unfamiliar enemy. Invisible to the naked eye, this enemy was more destructive than any the Romans had ever encountered in their long history. We cannot yet say with certainty what this microorganism was, although most historians strongly suspect that it was the debut of the smallpox virus, Variola major. (Hopefully, in coming years the pathogen’s DNA will be recovered from an archaeological sample, allowing a definite identification). In the meantime, the best testimony is furnished by the prolific medical writer, Galen, who lived through the pestilence and described a disease characterized by a ghastly pustular rash wrapping the entire body, a drawn out course of infection, hemorrhagic cases, and high mortality. There are simply few pathogens that are capable of accomplishing what this germ did. Within a few years, the disease rampaged from Egypt to Britain, from the Danube to the Sahara. The ancient witnesses report horrific mortality, and while accurate numbers are elusive, it is not unreasonable to think that the pestilence – known as the Antonine Plague – carried off 7 or 8 million victims in the Roman Empire. It was the single worst mortality event, in absolute terms, up to that point in human history.
The Antonine Plague struck the Roman Empire at its very apex. On the eve of the pestilence, the Roman Empire was the most powerful state in the world. Roman rule stretched from the shores of the Atlantic to the hills of Syria, from beyond the Danube to the first cataract of the Nile. One in four humans alive inhabited the empire’s borders. They enjoyed a rising standard of living, right down to the outbreak of the pestilence. The great mortality was no Malthusian meltdown. It was a bolt from the blue, a sharp bend in the course of history, triggered by the appearance of an emerging infectious disease from beyond the frontiers. The Antonine Plague was a demographic catastrophe. It did not topple the Roman Empire, nor did it condemn the empire to inevitable doom. But it did mark a turning point, a transition to an age characterized by sharper social and political challenges. In short, the mighty Roman Empire was transformed by a microscopic germ.
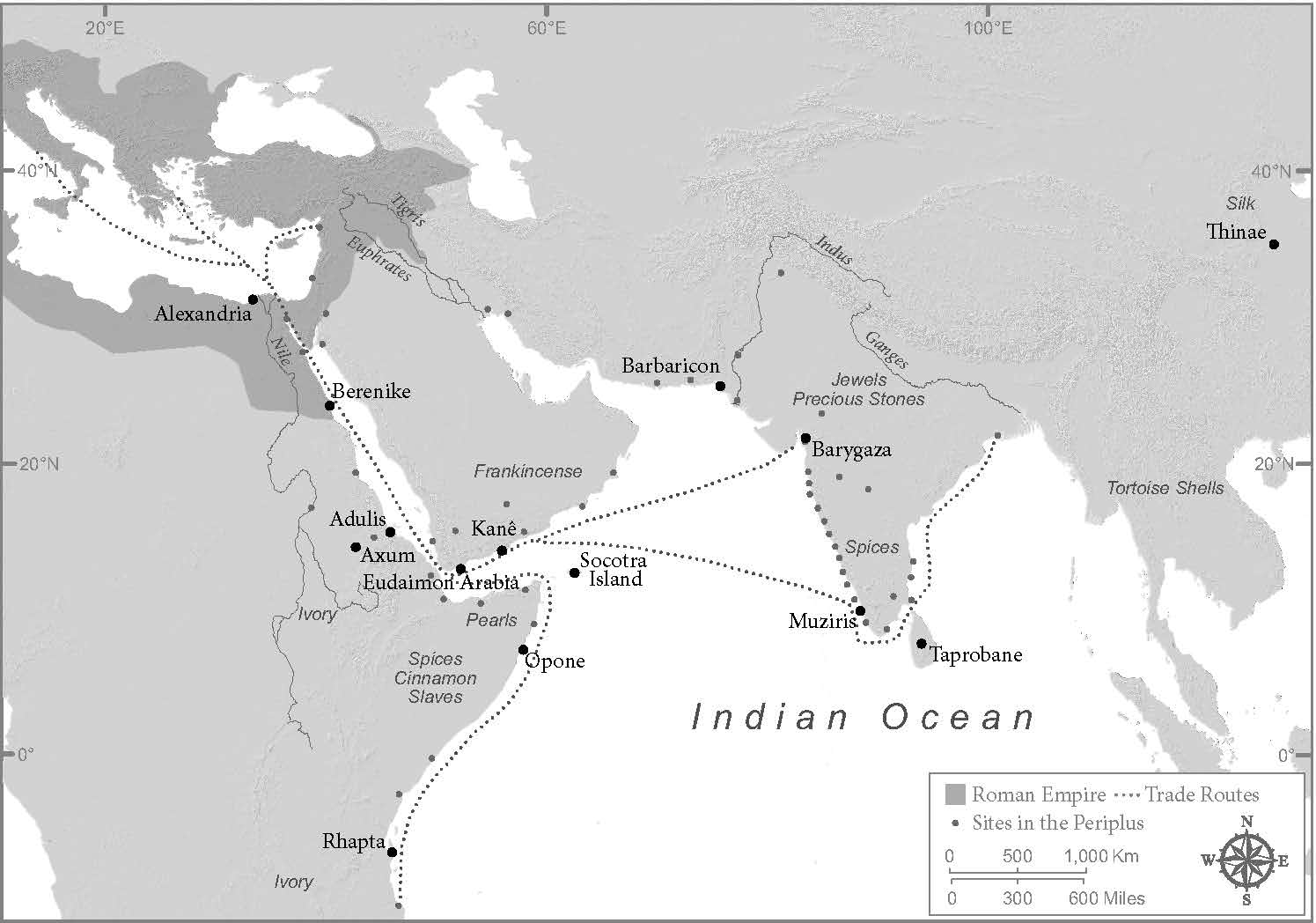
How are we to understand a biological event of this magnitude, which might deserve to be considered history’s first true “pandemic”? The answer, I would suggest, takes us to the heart of what Michael Gillings and Ian Paulsen have called “the microbiology of the Anthropocene.” The Antonine Plague occurred at the intersection of human and natural factors. If the disease was indeed smallpox, the agent of the pestilence evolved, relatively recently, in Africa, from an ancestral rodent Orthopoxvirus, and entered the Roman Empire as an evolutionary newborn. It was carried into the empire on the trading networks that had grown up in the first centuries, connecting the lands lying around the Indian Ocean. The Red Sea was a major theater of Roman trade, bringing spices, silks, ivories, slaves, etc. into the empire, in massive quantities. This trading network, the scene of incipient globalization, also brought germs. The Antonine Plague, then, was the conjuncture of human transformation of the planet with random, evolutionary events far beyond human influence.
Gillings and Paulsen show how we might apply the concept of the Anthropocene to the study of microbial organisms. They propose a framework in which human transformation of the earth’s systems has come to dominate the ecological and evolutionary conditions of microbial life. Gillings and Paulsen are concerned with the effects of human civilization on all microbial species, and they catalogue six principal ways in which human alteration of the environment has impacted bacteria, viruses, and protozoa: (1) the reconfiguration of the human microbiome, due principally to radical changes in the human diet; (2) the spread of antibiotic resistance, an evolutionary response to the widespread use of pharmaceuticals; (3) the transformation of global geochemical systems, including nitrogen and carbon cycles; (4) dispersal and disease (of most immediate interest here); (5) global warming and ocean acidification; and (6) the intentional human manipulation of microbial DNA. Gillings and Paulsen propose a timeline of human interference in which the rise of agriculture marks the beginnings of a Paleoanthropocene, the Industrial Revolution stands at the threshold of the Anthropocene proper, and the last 50 years should be considered the Great Acceleration.
In this post and one to follow, I’d like to explore the smaller subset of microorganisms that are capable of causing disease in humans. There are maybe a trillion microbial species on earth, but most of this wondrously diverse array is indifferent to us. Only something like 1400 species are known (so far) to be pathogenic to humans (over 500 bacteria, some 200 viruses, and assorted other fungi, helminths, and protozoa). These organisms in general possess the molecular weapons – virulence factors – that allow them to evade, subvert, or overcome our powerful immune systems. The background conditions of the physical climate shape and constrain the geographical ecology of pathogenic microbes, but human occupation of the globe has interacted with this natural background to configure the global landscape of germs that exists today. In the largest sense, the global distribution and prevalence of these pathogens is incomprehensible without accounting for human colonization of the planet and our transformation of planetary ecosystems.

World Health Organization Infectious and parasitic diseases world map
Consider, for instance, the WHO’s gargantuan efforts to catalogue the global distribution of infectious disease risk. The assessment immediately reveals that as far as dangerous microbes are concerned, it is not true that “everything is everywhere.” Many species are ecologically specific, and in particular the tropics bear a heavy burden of infectious disease. This pattern is due both to natural factors (i.e., pathogens obey the “latitudinal species gradient,” the greater diversity and richness of all life near the equator) and contingent historical factors (the strong association between underdevelopment and infectious disease risk). You are unlikely to die of falciparum malaria in Montreal. But the regional distribution of pathogens today also reflects the heavy hand of human intervention. Even in the few decades that the WHO has been trying to track these data, some germs have receded in importance (in fact, malaria, in many places) while other germs have emerged (for instance, the Zika virus), both of which are inconceivable without intentional and unintentional human manipulation of the environment. The WHO’s “map” of infectious disease is a product of both nature and human-driven ecological change.
From a germ’s point of view, human transformation of the evolutionary environment extends back millennia. To assess the human impact on the prevalence and diversity of infectious disease, we have to imagine the various mechanisms by which human transformation of the environment has altered the selection pressures operative for pathogenic microbes. In the words of the historian John McNeill, “for all species, on land and sea, the Anthropocene has revised the rules of evolution. Biological fitness— defined as success in the business of survival and reproduction— has increasingly hinged on compatibility with human enterprise. Those species that fit neatly into a humanized planet, such as pigeons, squirrels, rats, cattle, goats, crabgrass, rice, and maize prosper.” The same is manifestly true of pathogenic bacteria, viruses, protozoa, and other parasites. In Part II, we will classify some of the ways that our species has fundamentally changed the rules of the game for pathogenic microbes.

I have a question, and I hope it is not too stupid. You say the rise of the pestilence in 166 was the first true pandemic. How can we be sure of that? Maybe earlier pandemics in other cultures were simply undetected? And are there other reports of less lethal outbreaks of diseases? The system of Roman roads must have facilitated that, too, right?
As a Canadian, I feel obliged to point out that we did have severe malaria in Canada in several points in our history, and there is serious concern about its return (http://www.cmaj.ca/content/160/2/211.full.pdf). Although I suspect the disease itself was mostly imported (particularly by English soldiers who had spent time in the tropics), we do have large mosquito populations (and we often joke the mosquito should be our national bird!).
The broader point, however, is that patterns of biogeography are still poorly understood. Several groups of species have inverse latitudinal richness gradients (https://www.natur.cuni.cz/fakulta/zivotni-prostredi/lide/zamestnanci/pavel-kindlmann/dokumenty/inverse-20latitudinal-20gradients-20in-20species-20diversity.pdf as well as my own study: http://onlinelibrary.wiley.com/doi/10.1890/14-0717.1/full), and there are certainly continual arguments about underlying mechanisms (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4251432/). And so I find your point about interactions between human history and natural patterns of biogeography fascinating–how do we untangle the web of causality to get somewhere where we can be predictive?
Pingback: #EnvHist Worth Reading: October 2017 – NiCHE